



 i sposoby ich łagodzenia")
 i sposoby ich łagodzenia")






Glukagon to hormon peptydowy wytwarzany przez komórki alfa trzustki. Glukagon zwiększa stężenie glukozy i kwasów tłuszczowych we krwi i jest uważany za główny hormon kataboliczny organizmu. Glukagon jest również stosowany jako lek do leczenia wielu schorzeń. Działanie glukagonu jest odwrotne do działania insuliny, która obniża poziom glukozy pozakomórkowej. Glukagon jest produkowany z proglukagonu, kodowanego przez gen GCG.
Trzustka uwalnia glukagon, gdy ilość glukozy w krwiobiegu jest zbyt niska. Glukagon powoduje, że wątroba angażuje się w glikogenolizę: przekształcanie zmagazynowanego glikogenu w glukozę, która jest uwalniana do krwiobiegu. Z drugiej strony wysoki poziom glukozy we krwi stymuluje uwalnianie insuliny. Insulina umożliwia wychwyt glukozy i jej wykorzystanie przez tkanki insulinozależne. Tak więc glukagon i insulina są częścią systemu sprzężenia zwrotnego, który utrzymuje stabilny poziom glukozy we krwi. Glukagon zwiększa wydatek energetyczny i jest podwyższony w warunkach stresu. Glukagon należy do sekretynowej rodziny hormonów.
Funkcja glukagonu
Glukagon ogólnie podnosi stężenie glukozy we krwi poprzez promowanie glukoneogenezy i glikogenolizy. Glukagon zmniejsza również syntezę kwasów tłuszczowych w tkance tłuszczowej i wątrobie. Glukagon promuje lipolizę w tych tkankach, co powoduje, że uwalniają one kwasy tłuszczowe do krążenia, gdzie mogą być katabolizowane w celu wytworzenia energii w tkankach, takich jak mięśnie szkieletowe, gdy jest to wymagane.
Glukoza jest magazynowana w wątrobie w postaci glikogenu polisacharydowego, który jest glukanem (polimerem zbudowanym z cząsteczek glukozy). Komórki wątroby (hepatocyty) mają receptory glukagonu. Kiedy glukagon wiąże się z receptorami glukagonu, komórki wątroby przekształcają glikogen w pojedyncze cząsteczki glukozy i uwalniają je do krwiobiegu w procesie znanym jako glikogenoliza. Kiedy magazynowanie glukozy zostaje wyczerpane, glukagon zachęca wątrobę i nerki do syntezy dodatkowej glukozy przez glukoneogenezę. Glukagon wyłącza glikolizę w wątrobie, powodując przeniesienie pośrednich glikolitycznych związków do glukoneogenezy.
Glukagon reguluje również tempo produkcji glukozy poprzez lipolizę. Glukagon indukuje lipolizę u ludzi w warunkach supresji insuliny (takich jak cukrzyca typu 1).
Wydaje się, że produkcja glukagonu zależy od ośrodkowego układu nerwowego poprzez szlaki, które nie zostały jeszcze zdefiniowane. U bezkręgowców stwierdzono, że usuwanie szypułek ocznych wpływa na produkcję glukagonu. Wycięcie szypułki ocznej u młodych raków powoduje hiperglikemię wywołaną glukagonem.
Mechanizm działania glukagonu

Glukagon wiąże się z receptorem glukagonu, receptorem sprzężonym z białkiem G, zlokalizowanym w błonie komórkowej komórki. Zmiana konformacji receptora aktywuje białka G, heterotrimeryczne białko z podjednostkami α, β i γ. Kiedy białko G oddziałuje z receptorem, ulega zmianie konformacyjnej, która powoduje zastąpienie cząsteczki GDP, która była związana z podjednostką α, cząsteczką GTP. To podstawienie powoduje uwolnienie podjednostki α z podjednostek β i γ. Podjednostka alfa specyficznie aktywuje kolejny enzym w kaskadzie, cyklazę adenylanową.
Cyklaza adenylanowa wytwarza cykliczny monofosforan adenozyny (cykliczny AMP lub cAMP), który aktywuje kinazę białkową A (kinaza białkowa zależna od cAMP). Enzym ten z kolei aktywuje kinazę fosforylazy, która następnie fosforyluje fosforylazę glikogenu b (PYG b), przekształcając ją w aktywną formę zwaną fosforylazą a (PYG a). Fosforylaza jest enzymem odpowiedzialnym za uwalnianie glukozo-1-fosforanu z polimerów glikogenu.
Przykładem tego szlaku może być wiązanie się glukagonu z białkiem transbłonowym. Białka transbłonowe oddziałują z Gɑβ𝛾. Gɑ oddziela się od Gβ𝛾 i oddziałuje z transbłonową cyklazą adenylylową białka. Cyklaza adenylylowa katalizuje konwersję ATP do cAMP. cAMP wiąże się z kinazą białkową A, a kompleks fosforyluje kinazę fosforylazy. Fosforylowana kinaza fosforylazy fosforyluje fosforylazę. Fosforylowana fosforylaza odcina jednostki glukozy od glikogenu jako glukozo-1-fosforan.
Dodatkowo skoordynowana kontrola glikolizy i glukoneogenezy w wątrobie jest regulowana przez stan fosforylacji enzymów, które katalizują tworzenie silnego aktywatora glikolizy zwanego 2,6-bisfosforanem fruktozy. Enzym kinaza białkowa A (PKA), która była stymulowana kaskadą zainicjowaną przez glukagon, będzie również fosforylować pojedynczą resztę serynową dwufunkcyjnego łańcucha polipeptydowego zawierającego zarówno enzymy 2,6-bisfosfatazy fruktozy, jak i fosfofruktokinazę-2. Ta fosforylacja kowalencyjna zainicjowana przez glukagon aktywuje ten pierwszy i hamuje ten drugi. Reguluje to reakcję katalizującą 2,6-bisfosforan fruktozy (silny aktywator fosfofruktokinazy-1, enzymu będącego głównym etapem regulacji glikolizy) poprzez spowolnienie tempa jej powstawania, tym samym hamując przepływ szlaku glikolizy i umożliwiając glukoneogenezę dominować. Proces ten jest odwracalny w przypadku braku glukagonu (a tym samym obecności insuliny).
Stymulacja PKA glukagonem dezaktywuje również enzym glikolityczny kinazę pirogronianową w hepatocytach.
Fizjologia
Produkcja glukagonu
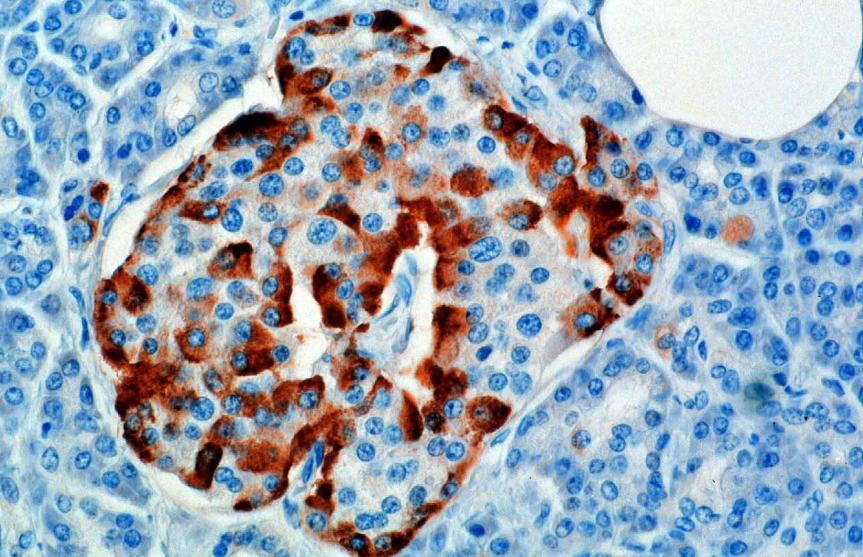
Hormon jest syntetyzowany i wydzielany z komórek alfa (komórek α) wysepek Langerhansa, które znajdują się w części dokrewnej trzustki. Produkcja, która skądinąd przebiega swobodnie, jest tłumiona/regulowana przez amylinę, hormon peptydowy wydzielany wspólnie z insuliną z komórek β trzustki. W miarę zmniejszania się poziomu glukozy w osoczu, późniejsze zmniejszenie wydzielania amyliny łagodzi jej tłumienie komórek α, umożliwiając wydzielanie glukagonu.
U gryzoni komórki alfa znajdują się na zewnętrznej krawędzi wysepki. Struktura ludzkiej wysepki jest znacznie mniej segregowana, a komórki alfa są rozmieszczone w całej wysepce w bliskim sąsiedztwie komórek beta. Glukagon jest również produkowany przez komórki alfa w żołądku.
Ostatnie badania wykazały, że produkcja glukagonu może również odbywać się poza trzustką, przy czym jelito jest najbardziej prawdopodobnym miejscem pozatrzustkowej syntezy glukagonu.
Regulacja glukagonu
Wydzielanie glukagonu jest stymulowane przez:
- Hipoglikemia
- Epinefryna (poprzez receptory adrenergiczne β2, α2 i α1)
- Arginina
- Alanina (często z transaminacji pirogronianu/glutaminianu pochodzącego z mięśni)
- Acetylocholina
- Cholecystokinina
- Polipeptyd hamujący żołądek
Wydzielanie glukagonu hamują:
- Somatostatyna
- Amylina
- Insulina (przez GABA)
- Heterodimer receptora PPARy/retinoidu X.
- Wzrost wolnych kwasów tłuszczowych i ketokwasów do krwi.
- Zwiększona produkcja mocznika
- Peptyd glukagonopodobny-1
Struktura glukagonu
Glukagon to 29-aminokwasowy polipeptyd. Jego pierwotna struktura u ludzi to: NH2-His-Ser-Gln-Gly-Thr-Phe-Thr-Ser-Asp-Tyr-Ser-Lys-Tyr-Leu-Asp-Ser-Arg-Arg-Ala-Gln-Asp-Phe-Val-Gln-Trp -Leu-Met-Asn-Thr-COOH.
Polipeptyd ma masę cząsteczkową 3485 daltonów. Glukagon jest hormonem peptydowym (niesteroidowym).
Glukagon jest generowany z rozszczepienia proglukagonu przez konwertazę proproteinową 2 w komórkach α wysepek trzustkowych. W jelitowych komórkach L proglukagon jest rozszczepiany na alternatywne produkty: glicentynę, GLP-1 (inkretyna), IP-2 i GLP-2 (wspomaga wzrost jelit).
Patologia
Nienormalnie podwyższone poziomy glukagonu mogą być spowodowane przez guzy trzustki, takie jak glukagonoma, których objawy obejmują nekrolityczny rumień migracyjny, zmniejszoną ilość aminokwasów i hiperglikemię. Może wystąpić samodzielnie lub w kontekście mnogiej gruczolakowatości wewnątrzwydzielniczej typu 1.
Podwyższony poziom glukagonu jest głównym czynnikiem przyczyniającym się do hiperglikemicznej kwasicy ketonowej w nierozpoznanej lub słabo leczonej cukrzycy typu 1. Ponieważ komórki beta przestają funkcjonować, insulina i GABA trzustki nie są już obecne, aby hamować uwalnianie glukagonu. W rezultacie glukagon jest maksymalnie uwalniany z komórek alfa, powodując szybki rozkład glikogenu do glukozy i szybką ketogenezę. Stwierdzono, że podgrupa dorosłych z cukrzycą typu 1 potrzebowała średnio 4 razy więcej czasu, aby zbliżyć się do kwasicy ketonowej, gdy podawano somatostatynę (hamuje produkcję glukagonu) bez insuliny. Hamowanie glukagonu jest popularnym pomysłem leczenia cukrzycy, jednak niektórzy ostrzegają, że może to spowodować kruchą cukrzycę u pacjentów z odpowiednio stabilnym poziomem glukozy we krwi.
Uważa się, że brak komórek alfa (a tym samym glukagonu) jest jednym z głównych czynników wpływających na skrajną zmienność poziomu glukozy we krwi w warunkach całkowitej pankreatektomii.
Historia
W latach dwudziestych Kimball i Murlin badali ekstrakty trzustkowe i znaleźli dodatkową substancję o właściwościach hiperglikemicznych. Opisali glukagon w 1923 roku. Sekwencja aminokwasowa glukagonu została opisana pod koniec lat pięćdziesiątych. Pełniejsze zrozumienie jego roli w fizjologii i chorobach zostało osiągnięte dopiero w latach 70., kiedy opracowano specjalny test radioimmunologiczny.
.
:max_bytes(150000):strip_icc()/GettyImages-150666718-579a41003df78c32762c6e54.jpg "10 wskazówek dotyczących gotowania dla osób z zapaleniem stawów")
Discussion about this post